
Elektrofiziológia - Electrophysiology
Az elektrofiziológia ( görögül ἥλεκτρον , ēlektron , "borostyán" [lásd az "elektron" etimológiáját ]; φύσις , physis , "természet, eredet"; és -λογία , -logia ) a fiziológia egyik ága, amely a biológiai elektromos tulajdonságait tanulmányozza sejtek és szövetek. Ez magában foglalja a feszültségváltozások vagy az elektromos áram mérését, vagy manipulációkat a legkülönfélébb skálákon, az egyetlen ioncsatornás fehérjékről az egész szervekre, például a szívre . Az idegtudományban magában foglalja a neuronok elektromos aktivitásának mérését , és különösen az akciós potenciál aktivitását. Az idegrendszerből származó nagyméretű elektromos jelek , például az elektroencefalográfia felvételeit elektrofiziológiai felvételeknek is nevezhetjük. Hasznosak az elektrodiagnosztika és a monitorozás során .

Meghatározás és hatály
Klasszikus elektrofiziológiai technikák
Elv és mechanizmusok
Az elektrofiziológia a fiziológia azon ága, amely széles körben vonatkozik az ionok áramlására ( ionáram ) a biológiai szövetekben, és különösen az elektromos rögzítési technikákra, amelyek lehetővé teszik ezen áramlás mérését. A klasszikus elektrofiziológiai technikák során elektródákat helyeznek el a biológiai szövetek különböző készítményeiben. Az elektródák fő típusai:
- egyszerű tömör vezetők, például tárcsák és tűk (egyes vagy tömbök, gyakran szigetelve, kivéve a csúcsot),
- nyomok nyomtatott áramköri lapokon vagy rugalmas polimereken, szintén szigetelve, kivéve a csúcsot, és
- elektrolittal töltött üreges csövek, például kálium -klorid -oldattal vagy más elektrolit -oldattal töltött üvegpipetták .
A fő előkészületek a következők:
- élő szervezetek (például rovaroknál ),
- kivágott szövet (akut vagy tenyésztett),
- disszociált sejtek a kivágott szövetből (akut vagy tenyésztett),
- mesterségesen tenyésztett sejtek vagy szövetek, ill
- a fentiek hibridjei.
A neuronok elektrofiziológiája az idegrendszeren belüli biológiai sejtek és szövetek elektromos tulajdonságainak tanulmányozása. Az idegsejtek elektrofiziológiájával az orvosok és a szakemberek az egyén agyi aktivitását vizsgálva megállapíthatják, hogyan történnek a neuronális rendellenességek. Olyan tevékenység, mint például az agy mely részei világítanak bármilyen helyzetben. Ha egy elektróda átmérője elég kicsi (mikrométer), akkor az elektrofiziológus dönthet úgy, hogy a csúcsot egyetlen cellába helyezi. Egy ilyen konfiguráció lehetővé teszi a közvetlen megfigyelés és intracelluláris felvétel a intracelluláris elektromos aktivitását egyetlen sejt. Ez az invazív beállítás azonban csökkenti a sejt élettartamát, és anyagszivárgást okoz a sejtmembránon. Az intracelluláris aktivitás elektrolitot tartalmazó, speciálisan kialakított (üreges) üvegpipettával is megfigyelhető. Ebben a technikában a mikroszkopikus pipettahegyet a sejtmembránhoz nyomják, amelyhez az üveg és a sejtmembrán lipidjei közötti kölcsönhatás révén szorosan tapad. A pipettán belüli elektrolit folyadékfolytonosságba hozható a citoplazmával úgy, hogy negatív nyomású impulzust juttat a pipettára annak érdekében, hogy eltörje a pipetta pereme által körülvett kis membránfoltot ( teljes sejt felvétel ). Alternatív megoldásként az ionos folytonosságot a tapasz "perforálásával" lehet létrehozni, lehetővé téve, hogy az elektroliton belüli exogén pórusképző szer beilleszkedjen a membránfoltba ( perforált tapasz rögzítése ). Végül a tapasz érintetlenül maradhat ( patch rögzítés ).
Az elektrofiziológus dönthet úgy, hogy nem helyezi a csúcsot egyetlen cellába. Ehelyett az elektróda csúcsa folytonosságban maradhat az extracelluláris térrel. Ha a hegy elég kicsi, ilyen konfiguráció lehetővé teszi a közvetett figyelése és rögzítése akciós potenciálok , egyetlen sejtből, nevezzük az egyetlen egységet felvétel . Attól függően, hogy a készítmény és precíz, extracelluláris konfiguráció vegye fel a tevékenységét több közeli sejtek egyszerre nevezett többlakásos felvétel .
Az elektróda méretének növekedésével a felbontóképesség csökken. A nagyobb elektródák csak sok sejt nettó aktivitására érzékenyek, ezeket helyi mezőpotenciáloknak nevezik . A még nagyobb elektródák, például a szigetelés nélküli tűk és a felszíni elektródák, amelyeket a klinikai és sebészeti neurofiziológusok használnak, csak bizonyos típusú szinkronaktivitásra érzékenyek a milliós számú sejtpopuláción belül.
Más klasszikus elektrofiziológiai technikák közé tartozik az egycsatornás rögzítés és az amperometria .
Elektrográfiai módszerek testrészenként
Az elektrofiziológiai felvételt általában néha elektrográfiának nevezik ( elektro- + -gráfia , "elektromos rögzítés"), az így előállított rekord elektrogram. Az elektrográfia szónak azonban más érzékei is vannak (beleértve az elektrofotográfiát is ), és az elektrofiziológiai rögzítés bizonyos típusait általában meghatározott nevekkel nevezik, amelyek az elektro- + [testrész kombináló forma ] + -grafika mintájára épülnek fel (ExG rövidítés). Hasonlóképpen, az elektrogram szó (nem szükséges más érzékszervek számára ) gyakran az intrakardiális elektrogramma sajátos jelentését hordozza, amely olyan, mint egy elektrokardiogram, de van néhány invazív vezetéke (a szív belsejében), nem pedig csak nem invazív vezetékek (a bőrön). A klinikai diagnosztikai célú elektrofiziológiai felvétel az elektrodiagnosztikai vizsgálatok kategóriájába tartozik . A különböző "ExG" módok a következők:
| Modalitás | Rövidítés | Testrész | Elterjedtség a klinikai alkalmazásban |
|---|---|---|---|
| elektrokardiográfia | EKG vagy EKG | szív (különösen a szívizom ), bőr elektródákkal (nem invazív) | 1 - nagyon gyakori |
| elektroatriográfia | EAG | pitvari szívizom | 3 - nem gyakori |
| elektroventrikulográfia | EVG | kamrai szívizom | 3 - nem gyakori |
| intrakardiális elektrogram | EGM | szív (különösen a szívizom ), intrakardiális elektródákkal (invazív) | 2 - kissé gyakori |
| elektroencefalográfia | EEG | agy (általában az agykéreg ), extrakraniális elektródákkal | 2 - kissé gyakori |
| elektrokortikográfia | EKG vagy iEEG | agy (különösen az agykéreg), koponyaűri elektródákkal | 2 - kissé gyakori |
| elektromiográfia | EMG | izmok az egész testben (általában csontvázak , néha simaak ) | 1 - nagyon gyakori |
| elektrookulográfia | EOG | szem - száz gömb | 2 - kissé gyakori |
| elektroretinográfia | ERG | szem - retina kifejezetten | 2 - kissé gyakori |
| elektronystagmográfia | HUN | szem - a corneoretinalis potenciálon keresztül | 2 - kissé gyakori |
| elektroolfactography | EOG | szaglóhám emlősökben | 3 - nem gyakori |
| elektroantennográfia | EAG | szagló receptorok ízeltlábú antennákban | 4 - klinikailag nem alkalmazható |
| elektrokochleográfia | ECOG vagy ECochG | belső fül | 2 - kissé gyakori |
| elektrogasztrográfia | TOJÁS | gyomor simaizom | 2 - kissé gyakori |
| elektrogasztroenterográfia | EGEG | a gyomor és a bél simaizomzatát | 2 - kissé gyakori |
| elektroglottográfia | TOJÁS | hangrés | 3 - nem gyakori |
| elektropalatográfia | EPG | nyelv palatális érintkezése | 3 - nem gyakori |
| elektroarteriográfia | EAG | artériás áramlás a bőrön keresztül észlelt áramlási potenciálon keresztül | 3 - nem gyakori |
| elektroblefalográfia | EBG | szemhéj izma | 3 - nem gyakori |
| elektrodermográfia | EDG | bőr | 3 - nem gyakori |
| elektrohysterography | EHG | méh | 3 - nem gyakori |
| elektroneuronográfia | ENeG vagy ENoG | idegek | 3 - nem gyakori |
| elektropneumográfia | EPG | tüdő (mellkasi mozgások) | 3 - nem gyakori |
| elektrospirográfia | ESG | gerincvelő | 3 - nem gyakori |
| elektrovomerográfia | EVG | vomeronasalis szerv | 3 - nem gyakori |
Optikai elektrofiziológiai technikák
Az optikai elektrofiziológiai technikákat tudósok és mérnökök hozták létre a klasszikus technikák egyik fő korlátjának leküzdésére. A klasszikus technikák lehetővé teszik az elektromos aktivitás megfigyelését a szövet térfogatának körülbelül egyetlen pontján. A klasszikus technikák szingularizálják az elosztott jelenséget. A bioelektromos aktivitás térbeli eloszlása iránti érdeklődés arra késztette a molekulákat, hogy elektromos vagy kémiai környezetükre fényt bocsássanak ki. Ilyenek például a feszültségérzékeny festékek és a fluoreszkáló fehérjék.
Miután egy vagy több ilyen vegyületet bejuttattak a szövetbe perfúzió, injekció vagy génexpresszió útján, megfigyelhető és rögzíthető az elektromos aktivitás 1 vagy 2 dimenziós eloszlása.
Intracelluláris felvétel
Az intracelluláris rögzítés magában foglalja a feszültség és/vagy áram mérését a cella membránján. Intracelluláris felvétel készítéséhez a finom (éles) mikroelektród hegyét be kell helyezni a cellába, hogy a membránpotenciál mérhető legyen. Általában egy egészséges sejt nyugalmi membránpotenciálja -60 és -80 mV között van, és az akciós potenciál alatt a membránpotenciál elérheti a +40 mV -ot. 1963 -ban Alan Lloyd Hodgkin és Andrew Fielding Huxley elnyerték az élettani vagy orvostudományi Nobel -díjat azért, mert hozzájárultak az idegsejtekben fellépő akciópotenciálok kialakulásának mechanizmusainak megértéséhez. Kísérleteik intracelluláris felvételeket tartalmaztak az atlanti tintahal óriási axonjából ( Loligo pealei ), és a "feszültségbilincs" technika első alkalmazásai közé tartoztak. Ma az intracelluláris rögzítéshez használt mikroelektródák többsége üveg mikropipetta, csúcsátmérőjük <1 mikrométer, ellenállása több megohm. A mikropipettákat olyan oldattal töltjük meg, amely ionos összetételű, mint a sejt intracelluláris folyadéka. A pipettába helyezett klórozott ezüsthuzal elektromosan köti össze az elektrolitot az erősítővel és a jelfeldolgozó áramkörrel. Az elektróda által mért feszültséget összehasonlítjuk egy referenciaelektród feszültségével, általában egy ezüst-kloriddal bevont ezüst huzal, amely a cella körüli extracelluláris folyadékkal érintkezik. Általában minél kisebb az elektróda csúcsa, annál nagyobb az elektromos ellenállása , így az elektróda kompromisszum a méret (elég kicsi ahhoz, hogy egyetlen cellába behatoljon, minimális károsodást okozva a sejtben) és az ellenállás (elég alacsony ahhoz, hogy kis idegsejtek felismerhető az elektróda csúcsán lévő termikus zaj miatt).
Feszültségbilincs

A feszültségbilincs -technika lehetővé teszi a kísérletező számára, hogy "rögzítse" a cella potenciálját egy kiválasztott értéken. Ez lehetővé teszi annak mérését, hogy adott feszültségnél mennyi ionáram halad át a sejt membránján. Ez azért fontos, mert sok a ioncsatornák a membrán egy neuron feszültségfüggő ioncsatornák , amelyek kinyitják csak akkor, ha a membrán feszültség egy bizonyos tartományon belül. Az áram feszültségbilincs-mérését az teszi lehetővé, hogy a rögzítőelektród és a sejtmembrán feltöltődésekor áthaladó kapacitív áramok közel egyidejű digitális kivonása megváltoztatja a cella potenciálját.
Árambilincs
Az aktuális rögzítő technika rögzíti a membránpotenciált úgy, hogy áramot injektál a cellába a rögzítőelektródon keresztül. Ellentétben a feszültségfogó üzemmóddal, ahol a membránpotenciált a kísérletező által meghatározott szinten tartják, az "áramfogó" módban a membránpotenciál szabadon variálható, és az erősítő rögzíti a cella által generált feszültséget önmagában vagy stimuláció eredménye. Ezt a technikát annak tanulmányozására használják, hogy a cella hogyan reagál, amikor egy elektromos áram belép a cellába; ez fontos például annak megértéséhez, hogy az idegsejtek hogyan reagálnak a neurotranszmitterekre, amelyek a membrán ioncsatornák megnyitásával hatnak .
A legtöbb áram-szorító erősítő alig vagy egyáltalán nem erősíti a cella által rögzített feszültségváltozásokat. Az "erősítő" valójában egy elektrométer , amelyet néha "egység erősítő erősítőnek" is neveznek; fő célja, hogy csökkentse a cellák által keltett kis jelek elektromos terhelését (mV tartományban), hogy azokat alacsony impedanciájú elektronika pontosan rögzítse . Az erősítő növeli az áramot a jel mögött, miközben csökkenti az ellenállást, amely felett az áram áthalad. Tekintsük ezt a példát Ohm törvénye alapján: 10 mV feszültséget generálunk, ha 10 nanoamper áramot vezetünk át 1 MΩ ellenálláson. Az elektrométer ezt a "nagy impedanciájú jelet" "alacsony impedanciájú jellé" változtatja egy feszültségkövető áramkör használatával. A feszültségkövető leolvassa a bemeneten lévő feszültséget (amelyet egy kis ellenálláson keresztül okozott kis áram okoz ). Ezután utasít egy párhuzamos áramkört, amely mögött nagy áramforrás van (az elektromos hálózat), és beállítja a párhuzamos áramkör ellenállását, hogy ugyanaz a kimeneti feszültség, de kisebb ellenállás mellett.
Patch-clamp rögzítés

Ezt a technikát Erwin Neher és Bert Sakmann fejlesztették ki, akik 1991 -ben Nobel -díjat kaptak. A hagyományos intracelluláris felvétel magában foglalja a cella finom elektródával történő beültetését; A patch-clamp rögzítés más megközelítést alkalmaz. A patch-clamp mikroelektróda egy viszonylag nagy csúcsátmérőjű mikropipetta. A mikroelektródát egy cella mellé kell helyezni, és a mikroelektródán keresztül enyhe szívást kell végezni, hogy a sejtmembrán egy darabját (a „tapaszt”) a mikroelektród csúcsába vonják; az üveghegy nagy ellenállású „tömítést” képez a sejtmembránnal. Ez a konfiguráció a "cellához csatolt" mód, és felhasználható a membránfoltban lévő ioncsatornák aktivitásának tanulmányozására. Ha most nagyobb szívóerőt alkalmazunk, az elektróda csúcsán lévő kis membránfolt elmozdulhat, így az elektróda lezárva marad a cella többi részén. Ez az "egész cellás" mód nagyon stabil intracelluláris felvételt tesz lehetővé. Hátrány (összehasonlítva a hagyományos intracelluláris rögzítéssel, éles elektródákkal), hogy a cella intracelluláris folyadéka keveredik a rögzítőelektródon belüli oldattal, és így az intracelluláris folyadék néhány fontos összetevője felhígítható. Ennek a technikának egy változata, a "perforált folt" technika megpróbálja minimalizálni ezeket a problémákat. Ahelyett, hogy szívnánk a membránfoltot az elektróda hegyéről, kiszorítanánk, lehetőség van arra is, hogy apró lyukakat készítsünk a tapaszon pórusképző szerekkel, így nagy molekulák, például fehérjék maradhatnak a sejt belsejében, és az ionok szabadon átjuthatnak a lyukakon . A membránfolt is eltávolítható a sejt többi részétől. Ez a megközelítés lehetővé teszi a tapasz membrántulajdonságainak farmakológiai elemzését.
Éles elektróda rögzítés
Azokban a helyzetekben, amikor a sejtmembránon belüli potenciált akarjuk rögzíteni, minimális hatással az intracelluláris folyadék ionösszetételére, éles elektródát használhatunk. Ezek a mikropipetták (elektródák) ismét olyanok, mint az üvegkapillárisokból húzott tapaszbilincsek, de a pórusok sokkal kisebbek, így nagyon kevés az ioncsere az intracelluláris folyadék és a pipetta elektrolitja között. A mikropipetta elektróda elektromos ellenállását csökkenti, ha 2-4 M KCl-el töltjük meg, nem pedig sókoncentrációval, amely utánozza a tapasz rögzítésénél alkalmazott intracelluláris ionkoncentrációt. Gyakran az elektróda hegye tele van különféle festékekkel, például Lucifer sárgával, hogy kitöltse a rögzített sejteket, morfológiájuk mikroszkóp alatt történő megerősítésére. A festékeket úgy fecskendezik be, hogy pozitív vagy negatív, egyenáramú vagy impulzusos feszültséget alkalmaznak az elektródákon, a festék polaritásától függően.
Extracelluláris felvétel
Egyegységes felvétel
Egy élő állat agyába bejuttatott elektróda észleli az elektromos aktivitást, amelyet az elektróda csúcsa melletti neuronok generálnak. Ha az elektróda mikroelektróda, csúcsa körülbelül 1 mikrométer, akkor az elektróda általában legfeljebb egy neuron aktivitását érzékeli. Az ilyen módon történő rögzítést általában "egyegységes" felvételnek nevezik. A rögzített akciós potenciálok nagyon hasonlítanak az intracellulárisan rögzített akciós potenciálokhoz, de a jelek sokkal kisebbek (jellemzően körülbelül 1 mV). A legtöbb felvétel az egyes idegsejtek aktivitásáról altatott és tudatos állatokban ilyen módon készül. Az élő állatok egyetlen neuronjának felvételei fontos betekintést nyújtottak abba, hogy az agy hogyan dolgozza fel az információkat. Például David Hubel és Torsten Wiesel rögzítették az egyes idegsejtek aktivitását az altatott macska elsődleges látókéregében , és bemutatták, hogyan reagálnak ezen a területen az egyes idegsejtek a vizuális inger nagyon sajátos jellemzőire. Hubel és Wiesel 1981 -ben fiziológiai vagy orvosi Nobel -díjat kapott.
Több egység rögzítése
Ha az elektróda csúcsa valamivel nagyobb, akkor az elektróda rögzítheti a több neuron által generált aktivitást. Az ilyen típusú felvételt gyakran "több egységből álló rögzítésnek" nevezik, és gyakran használják tudatos állatoknál, hogy rögzítsék az aktivitás változásait egy különálló agyterületen normál tevékenység során. Egy vagy több ilyen, egymástól közeli elektródáról készített felvételek segítségével azonosítani lehet a körülötte lévő cellák számát, valamint azt, hogy a tüskék melyik cellából származnak. Ezt a folyamatot tüskés szortírozásnak nevezik, és alkalmas olyan területeken, ahol azonosított sejttípusok vannak, amelyek jól meghatározott tüskejellemzőkkel rendelkeznek. Ha az elektróda csúcsa még mindig nagyobb, általában nem lehet megkülönböztetni az egyes neuronok aktivitását, de az elektróda továbbra is képes rögzíteni a sok sejt aktivitása által generált mezőpotenciált.
Terepi potenciálok

Az extracelluláris mezőpotenciálok helyi áramelnyelők vagy források, amelyeket sok sejt kollektív tevékenysége generál. Általában egy mezőpotenciált sok neuron egyidejű aktiválása generál szinaptikus átvitel útján . A jobb oldali diagram a hippokampális szinaptikus mező potenciáljait mutatja. A jobb oldalon az alsó nyom negatív negatív hullámot mutat, amely megfelel a jelenlegi elnyelésnek, amelyet a pozitív töltések okoznak a sejtekbe a posztszinaptikus glutamátreceptorokon keresztül , míg a felső nyoma pozitív hullámot mutat, amelyet a sejtből (a sejtnél) távozó áram generál test) az áramkör befejezéséhez. További információkért tekintse meg a helyi terepi potenciált .
Amperometria
Az amperometria szénelektródával rögzíti a biológiai oldat oxidált összetevőinek kémiai összetételében bekövetkezett változásokat. Az oxidációt és a redukciót a rögzítő elektróda aktív felületén lévő feszültség megváltoztatásával érik el a "szkennelés" néven ismert folyamatban. Mivel bizonyos agyi vegyi anyagok jellegzetes feszültségen elektronokat veszítenek vagy nyernek, az egyes fajok azonosíthatók. Az amperometriát az exocitózis tanulmányozására használták az idegrendszerben és az endokrin rendszerben. Sok monoamin neurotranszmitter ; pl. a noradrenalin (noradrenalin), a dopamin és a szerotonin (5-HT) oxidálhatók. A módszer alkalmazható olyan sejtekkel is, amelyek nem választanak ki oxidálható neurotranszmittereket, ha 5-HT-vel vagy dopaminnal "töltik fel" azokat.
Sík patch bilincs
A Planar patch clamp egy új módszer, amelyet nagy teljesítményű elektrofiziológiára fejlesztettek ki. Ahelyett, hogy pipettát helyezne egy tapadó cellára, a sejtszuszpenziót egy mikrostrukturált nyílást tartalmazó chipre pipettázzuk . Ezután egyetlen cellát helyeznek el a lyukon szívással, és szoros kapcsolatot (Gigaseal) alakítanak ki. A síkgeometria számos előnyt kínál a klasszikus kísérlethez képest:
- Lehetővé teszi a mikrofluidikák integrálását , ami lehetővé teszi a vegyületek automatikus alkalmazását az ioncsatorna -szűréshez.
- A rendszer hozzáférhető optikai vagy szkennelési szondatechnikákhoz .
- Az intracelluláris oldal perfúziója elvégezhető.
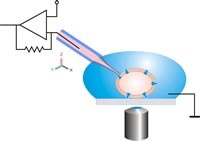
A klasszikus patch bilincs konfiguráció sematikus rajza. A tapasz -pipettát mikromanipulátor segítségével optikai vezérléssel mozgatják a cellába. Kerülni kell a pipetta és a cella közötti relatív mozgásokat, hogy a cella-pipetta kapcsolat sértetlen maradjon.

Tapasz pipetta szkennelő elektronmikroszkóp képe.
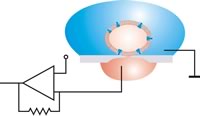
Sík foltkonfigurációban a cella szívással helyezkedik el. A zárás után a záróelem és a rekesz közötti relatív mozgások kizárhatók. A rezgéscsillapító tábla nem szükséges.

Lapos patch bilincs chip pásztázó elektronmikroszkóp képe. A pipetta és a chip is boroszilikát üvegből készült.




Más módszerek
Szilárdan támogatott membrán (SSM) alapú
Ezzel elektrofiziológiai megközelítéssel, proteo liposzómák , membrán vezikulumok , vagy membrán fragmentumokat tartalmazó csatorna vagy transzporter érdekes adszorbeáltunk egy lipidegyréteg festett egy funkcionalizált elektród. Ez elektróda állhat egy pohár támogató, a króm réteget, egy arany réteget, és egy oktadecil -merkaptán egyrétegű. Mivel a festett membránt az elektróda támogatja, szilárd hordozós membránnak nevezik. Fontos megjegyezni, hogy a mechanikai zavarok, amelyek általában elpusztítják a biológiai lipid membránt, nem befolyásolják az SSM élettartamát. A kapacitív elektróda (amely az SSM -ből és az elnyelt vezikulákból áll) mechanikailag annyira stabil, hogy az oldatok gyorsan cserélhetők a felületén. Ez a tulajdonság lehetővé teszi a gyors szubsztrát/ligandum koncentráció ugrások alkalmazását az érdeklődésre számot tartó fehérje elektrogenikus aktivitásának vizsgálatához, amelyet a vezikulák és az elektróda közötti kapacitív kapcsoláson keresztül mérünk.
Bioelektromos felismerési vizsgálat (BERA)
A bioelektromos felismerési vizsgálat (BERA) egy új módszer különböző kémiai és biológiai molekulák meghatározására a gélmátrixban rögzített sejtek membránpotenciáljában bekövetkező változások mérésével. Az elektróda-cella interfész fokozott stabilitása mellett az immobilizáció megőrzi a sejtek életképességét és fiziológiai funkcióit. A BERA -t elsősorban bioszenzoros alkalmazásokban használják olyan analitok vizsgálatára, amelyek kölcsönhatásba léphetnek az immobilizált sejtekkel a sejtmembrán potenciáljának megváltoztatásával. Ily módon, ha pozitív mintát adnak az érzékelőhöz, az elektromos potenciál jellegzetes, "aláírásszerű" változása következik be. A BERA a nemrégiben elindított páneurópai FOODSCAN projekt alaptechnológiája, amely az európai peszticid- és élelmiszerkockázat-értékelésről szól. A BERA -t humán vírusok ( hepatitisz B és C vírusok és herpeszvírusok ), állatbetegségek ( száj- és körömfájás vírus, prionok és kéknyelv vírus ) és növényi vírusok (dohány- és uborka vírusok) kimutatására használták specifikus, gyors (1-2 perc), reprodukálható és költséghatékony módon. A módszert alkalmazták a környezeti toxinok, például az élelmiszerekben található peszticidek és mikotoxinok , valamint a parafa és a bor 2,4,6-triklór -anizoljának kimutatására , valamint a szuperoxid- anion nagyon alacsony koncentrációjának meghatározására a klinikai mintákban .
A BERA érzékelő két részből áll:
- A fogyasztható biofelismerő elemek
- Beépített mesterséges intelligenciával rendelkező elektronikus leolvasó eszköz .
A közelmúlt előrelépése a membrántechnológián (MIME) nevű molekuláris azonosításnak nevezett technika kifejlesztése. Ez a technika lehetővé teszi, hogy gyakorlatilag bármilyen érdeklődésre számot tartó molekulára meghatározott specifitással rendelkező sejteket építsenek, több ezer mesterséges receptor beágyazásával a sejtmembránba.
Számítási elektrofiziológia
Bár nem szigorúan kísérleti mérés, módszereket fejlesztettek ki a fehérjék és biomembránok in silico vezetőképességének vizsgálatára . Ezek elsősorban molekuláris dinamikai szimulációk, amelyekben egy olyan modellrendszert, mint egy lipid kettősréteg , külső feszültségnek tesznek ki. Az ezeket a beállításokat használó vizsgálatok olyan dinamikus jelenségeket tudtak tanulmányozni, mint a membránok elektroporációja és az ionok transzlokációja csatornákon keresztül.
Az ilyen módszerek előnye az aktív vezetési mechanizmus magas szintű részletessége, amelyet az atomisztikus szimuláció által biztosított eredendően magas felbontás és adatsűrűség ad. A modell legitimitásának bizonytalansága és a modellezési rendszerek számítási költségei miatt jelentős hátrányok vannak, amelyek elég nagyok és elegendő időtartamúak ahhoz, hogy a rendszerek makroszkopikus tulajdonságait reprodukálni lehessen. Míg az atomisztikus szimulációk elérhetik a mikroszekundumos tartományhoz közeli vagy abba tartozó időskálákat, ez még mindig nagyságrendekkel alacsonyabb, mint a kísérleti módszerek, például a patch-clamping felbontása.
Klinikai elektrofiziológia
A klinikai elektrofiziológia azt vizsgálja, hogyan alkalmazhatók az elektrofiziológiai elvek és technológiák az emberi egészségre. Például a klinikai szív elektrofiziológiája a szívritmust és aktivitást szabályozó elektromos tulajdonságok tanulmányozása. A szív elektrofiziológiája alkalmazható olyan rendellenességek megfigyelésére és kezelésére, mint az aritmia (szabálytalan szívverés). Például az orvos behelyezhet egy elektródát tartalmazó katétert a szívbe, hogy rögzítse a szívizom elektromos aktivitását.
A klinikai elektrofiziológia másik példája a klinikai neurofiziológia . Ebben az orvosi szakterületen az orvosok mérik az agy , a gerincvelő és az idegek elektromos tulajdonságait . Az olyan tudósok, mint Duchenne de Boulogne (1806–1875) és Nathaniel A. Buchwald (1924–2006), úgy vélik, hogy nagymértékben előrehaladtak a neurofiziológia területén , lehetővé téve annak klinikai alkalmazását.
Klinikai jelentések
A minimális információs (MI) szabványok vagy jelentési irányelvek meghatározzák azt a minimális mennyiségű metaadatot (információt) és adatot, amely egy adott cél vagy célok eléréséhez szükséges egy klinikai vizsgálat során. A "Minimális információ egy idegtudományi vizsgálatról" (MINI) jelentéstételi útmutató dokumentumok családja célja, hogy következetes iránymutatásokat nyújtson az elektrofiziológiai kísérlet jelentéséhez. A gyakorlatban a MINI modul tartalmaz egy ellenőrző listát azokról az információkról, amelyeket meg kell adni (például az alkalmazott protokollokról), amikor egy adatkészletet közzétételre írnak le.
Lásd még
- Automatizált patch bilincs
- Bioelektrokémia
- Bioelektromágneses
- A szív elektrofiziológiája
- Klinikai szív elektrofiziológia
- Klinikai elektrofiziológia
- Klinikai neurofiziológia
- Elektrofiziológiai tanulmány
- Hille -egyenlet
- Multiscale Electrophysiology Format
- Neurofiziológia
- Szelet előkészítése
- Transzkután elektromos ideg stimuláció
Hivatkozások
Külső linkek